

La sangre del draco es incolora porque carece de hemoglobina , la proteína que se une al oxígeno en la sangre. Channichthyidae son los únicos vertebrados conocidos que carecen de hemoglobina como adultos. Aunque no fabrican hemoglobina, se pueden encontrar restos de genes de hemoglobina en su genoma . La proteína de hemoglobina está compuesta de dos subunidades (alfa y beta). En 15 de las 17 especies de draco, el gen de la subunidad beta ha sido completamente eliminado y el gen de la subunidad alfa ha sido parcialmente eliminado. En solo una de las especies de draco rayado, Neopagetopsis ionah , existe un gen de hemoglobina más completo pero aún no funcional. Los glóbulos rojos (RBC) generalmente están ausentes, y si están presentes, son raros y difuntos.
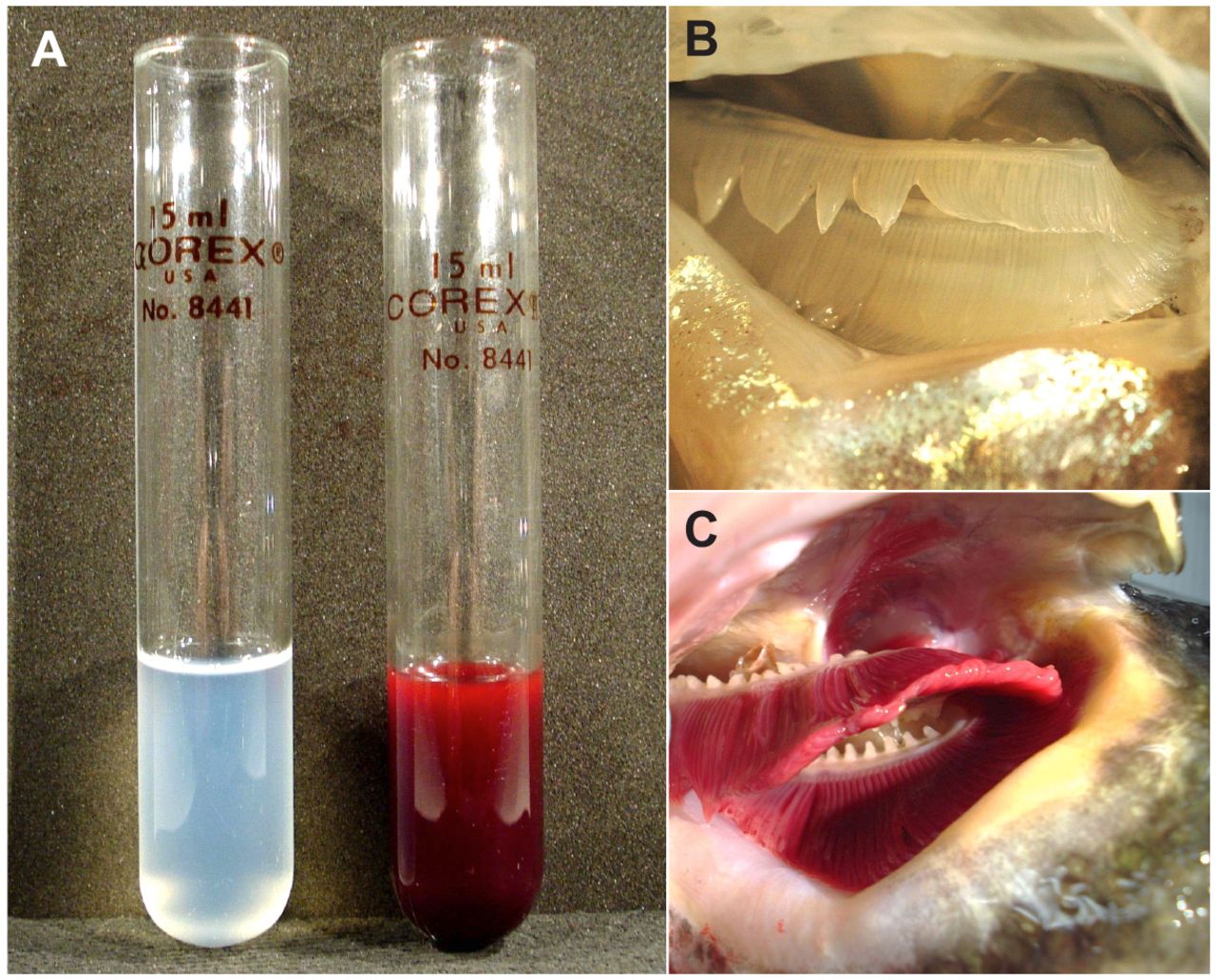

Para compensar la pérdida de hemoglobina, tienen vasos sanguíneos más grandes (incluidos los capilares ), mayores volúmenes de sangre (cuatro veces más que otros peces), corazones más grandes y un mayor gasto cardíaco (cinco veces mayor) en comparación con otros peces. Sus corazones carecen de arterias coronarias , y los músculos del ventrículo son muy esponjosos, lo que les permite absorber oxígeno directamente de la sangre que bombean. Sus corazones, vasos sanguíneos grandes y sangre de baja viscosidad (sin glóbulos rojos) están especializados para llevar a cabo caudales muy altos a bajas presiones. Esto ayuda a reducir los problemas causados por la falta de hemoglobina. En el pasado, se pensaba que su piel sin escamas ayudaba a absorber oxígeno. Sin embargo, el análisis actual ha demostrado que la cantidad de oxígeno absorbida por la piel es mucho menor que la absorbida por las branquias. El poco oxígeno extra absorbido por la piel puede desempeñar un papel en la suplementación del suministro de oxígeno al corazón que recibe sangre venosa de la piel y el cuerpo antes de bombearla a las branquias.

La pérdida de hemoglobina se pensó inicialmente como una adaptación al frío extremo ya que la falta de hemoglobina y de glóbulos rojos disminuye la viscosidad sanguínea, que es una adaptación que se ha observado en especies adaptadas a climas fríos. Sin embargo, el análisis actual ha demostrado que la falta de hemoglobina, aunque no es letal, no es adaptativa. Todas las ventajas adaptativas derivadas de la reducción de la viscosidad de la sangre se ven compensadas por el hecho de que los peces de hielo deben bombear mucha más sangre por unidad de tiempo para compensar la menor capacidad de transporte de oxígeno de su sangre. El alto volumen de sangre de los peces de hielo es evidencia de que la pérdida de hemoglobina y mioglobina no fue ventajosa para el ancestro de los peces de hielo. Su fisiología cardiovascular inusual, que incluye corazón grande, volumen sanguíneo alto, densidad mitocondrial aumentada y microvasculatura extensa, sugiere que los peces helados han tenido que desarrollar formas de lidiar con los daños a sus sistemas de transporte y atascamiento de oxígeno deteriorados.

Como se discutió previamente, las relaciones filogenéticas indican que la expresión de mioglobina en el tejido cardíaco se ha producido al menos cuatro veces. Esta pérdida repetida sugiere que la mioglobina cardíaca podría ser vestigial o incluso perjudicial para los peces de hielo. Sidell y O'Brien (2006) investigaron esta posibilidad. Primero, realizaron una prueba usando espectrometría de flujo detenido. Descubrieron que a través de todas las temperaturas, el oxígeno se une y se disocia más rápidamente del draco rayado que de la mioglobina de mamíferos. Sin embargo, cuando repetían la prueba con cada organismo a una temperatura que reflejaba con precisión su entorno nativo, el rendimiento de la mioglobina era más o menos equivalente entre los peces de hielo y los mamíferos.


La Antártida es un ambiente atípico. Para empezar, el Océano Austral se ha caracterizado por temperaturas extremadamente frías pero estables durante los últimos 10 a 14 millones de años. [15] Estas temperaturas frías, que permiten un mayor contenido de oxígeno en el agua, combinado con un alto grado de mezcla vertical en estas aguas, significa que hay una disponibilidad inusualmente alta de oxígeno en las aguas antárticas. Para un pez que ha perdido sus proteínas que se unen al oxígeno, esta es una propiedad muy fortuita. La pérdida de hemoglobina y mioglobina tendría consecuencias negativas en ambientes más cálidos. La estabilidad de la temperatura antes mencionada también es "afortunada", ya que las fuertes fluctuaciones de temperatura crearían un ambiente más estresante que probablemente eliminaría a los individuos con mutaciones perjudiciales. Los factores biológicos, en particular un nivel inusualmente bajo de competencia interespecífica, también contribuyeron a la fijación de estos rasgos inesperados. Parte de esto se debe a que hubo una grave caída en la biodiversidad de los peces entre mediados del Terciario y el presente. En segundo lugar, los factores físicos han contribuido al relativo aislamiento de los hielos, incluidas las corrientes circumpolares, las zanjas que rodean el continente antártico y, a lo largo de los últimos 5 millones de años, la presencia esporádica de refugios en forma de puertos marinos libres de hielo. Por lo tanto, estos rasgos lograron solucionarlo, no porque fueran adaptativos, sino porque las condiciones permitían que un rasgo de inadaptación no se seleccionara. Sin embargo, otros mecanismos compensatorios parecen haberse arreglado, lo que sugiere la presencia de alguna presión evolutiva. Esto crea una especie de paradoja.

La clave para resolver este enigma es considerar la otra función que desempeñan tanto la hemoglobina como la mioglobina. Si bien a menudo se hace hincapié y es comprensible sobre la importancia de la hemoglobina y la mioglobina en el suministro y el uso de oxígeno, estudios recientes han encontrado que ambas proteínas también están involucradas en el proceso de descomponer el óxido nítrico. Esto significa que cuando los peces helados perdieron hemoglobina y mioglobina no solo significó una menor capacidad para transportar oxígeno, sino que también aumentó el nivel total de óxido nítrico en las personas. El óxido nítrico desempeña un papel en la regulación de diversos procesos cardiovasculares en peces helados tales como la dilatación de la vasculatura branquial, el volumen del golpe cardíaco y la potencia de salida.


Estos peces han descendido de un ancestro demersal lento. Las aguas frías, bien mezcladas y ricas en oxígeno del Océano Austral proporcionaron un entorno en el que un pez con una baja tasa metabólica podría sobrevivir incluso sin hemoglobina, aunque de manera menos eficiente. Durante el período Terciario medio, una colisión de especies en el Océano Austral abrió una amplia gama de nichos vacíos para colonizar. A pesar de que los mutantes sin hemoglobina estaban menos en forma, la falta de competencia permitió incluso que los mutantes dejaran descendientes que colonizaran hábitats vacíos y compensaciones evolucionadas para sus mutaciones. Más tarde, las aperturas periódicas de los fiordos crearon hábitats que fueron colonizados por unos pocos individuos. Estas condiciones pueden haber permitido la pérdida de mioglobina. Actualmente se reconocen 17 especies de draco rayado agrupadas en 11 géneros:

Género Chaenocephalus (1913).- Es un género monotipico de la familia Channichthyidae del orden de los Perciformes.

Chaenocephalus aceratus (1906) - Draco antártico o Pez hielo austral.- Es una especie de draco rayado que se encuentra en la isla de Bouvet y en la península antártica del norte , donde se encuentra a una profundidad de 5 a 770 metros (16 a 2,526 pies). Esta especie crece a una longitud de 72 centímetros (28 in) LT. Longitud común 50 cm. Peso máximo publicado 3,7 kg. El draco rayado es aparentemente único entre los vertebrados ya que carece tanto de glóbulos rojos como de hemoglobina. Su única forma de transporte de oxígeno es oxígeno disuelto en su plasma sanguíneo. Hay solo una generación de ovocitos en los ovarios indicativos de fecundidad determinada. Las larvas tienen una larga fase pelágica. Las hembras maduras pueden reproducirse por primera vez alrededor de los 6-8 años de edad. Los adultos se alimentan principalmente de peces y krill. Generador síncrono. Utilizado como un pez de alimento. Es de menor importancia para las pesquerías comerciales. Se encuentra en el Océano Austral: en las cercanías de la isla de Bouvet, el mar de Scotia y la parte norte de la Península Antártica. Esta especie es el único miembro conocido de su género

Género Chaenodraco (1914).- Es un género monotipico de la familia Channichthyidae del orden de los Perciformes.

Chaenodraco wilsoni (1914) - Draco espinudo.- Es una especie de draco rayado nativo del Océano Austral, donde ocurre a profundidades de 200 a 800 metros (660 a 2.620 pies). Esta especie crece a una longitud de 43 centímetros (17 pulgadas ) de TL. Espinas dorsales 5 - 8; Radios dorsales blandos 38-42; Radios suaves anales 32 - 36. Cresta supraorbital no crenulada. Línea lateral media restringida al pedúnculo caudal; línea lateral inferior que se origina desde el origen del aleta anal hasta el sexto radio anterior anal. Maxilar que se extiende hasta el tercio anterior inferior del ojo. Huesos operculares con 2 espinas superiores y 3 inferiores. Canal preopercular-mandibular no unido al canal temporal. Aletas pélvicas que se extienden más allá del origen de la aleta anal. En la vida, gris pálido, blanquecino ventralmente. Diferenciado por la forma oval dorsoventralmente; el prominente rostro redondeado distalmente, pseudo-rostrum y pseudo-antirostrum; la excisura ostii y pseudo-excisura ostii claramente definidas; el coll agudamente estrecho; y el colliculli bien desarrollado. Ontogenia: La excisura ostii y la pseudo-excisura ostii se vuelven más profundas y la cresta inferior se vuelve menos distinta con un aumento en el tamaño del pez.

La fase pelágica larval es larga. La comida consiste de peces y krill. Engendro en invierno. Común en las aguas menos profundas de la plataforma continental, especialmente en los bancos de menos de 250 m de profundidad en áreas donde la afloración local aumenta el suministro de alimentos. Postlarvas y juveniles pelágicos también se encuentran en los 100 m superiores. Presa de pingüinos y focas. Utilizado como un pez comestible. Es de menor importancia para las pesquerías comerciales . Esta especie es el único miembro conocido de su género. Se encuentra en el Océano Austral: Circun-Antártico en la plataforma continental antártica. Registros más septentrionales de las proximidades de las Islas Orcadas del Sur y la Península Antártica.

Género Champsocephalus (1862).- Es un género de peces helados nativos del Océano Austral. Actualmente hay dos especies reconocidas en este género:

Champsocephalus esox (1861).- Es una especie de draco rayado que se encuentra a profundidades de 50-250 m (160-820 pies) en los océanos frente a la Patagonia , las Malvinas y Georgia del Sur. Alcanza los 35 cm (14 pulgadas) de longitud. Espinas dorsales 9; Radios blandos dorsales 32; Radios suaves anales 31; Vértebras 57-60. Margen posterior del emarginado de la aleta caudal. Sin escamas, excepto en la línea lateral. Cuerpo marrón, irregularmente moteado. Se alimenta principalmente de peces y krill. Se encuentra en el Océano Austral: región de Falkland Patagónica, Estrecho de Magallanes y raramente en Georgia del Sur.

Champsocephalus gunnari (1905) - Draco rayado o Pez hielo común.- Es un pez que se encuentra solo en el Océano Austral . Se encuentran principalmente cerca de las islas Heard y McDonald , Îles Kerguelen e islas en el Atlántico sur , como Georgia del Sur. Su tamaño normal es 22-25 pulgadas de largo, y solo 1 pulgada cuando nace. Vértebras 59 - 62. Se reproduce en otoño e invierno. Generador síncrono. La fase pelágica larval es larga. Se alimenta principalmente de krill y mysids. Utilizado como un pez de alimento. Debido a la sobrepesca, la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) considera que la especie está "agotada". Se encuentra en el Océano Austral: Islas del Mar de Scotia, incluida la parte norte de la Península Antártica; Kerguelen, Heard y las islas Bouvet.

Género Channichthys (1844).- Es un género de la familia Channichthyidae del orden de los Perciformes nativos del Océano Austral. Actualmente hay diez especies reconocidas en este género:

Channichthys aelitae (1995).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Los machos pueden alcanzar los 33,4 cm de longitud máxima. Se encuentra en el Océano meridional: Meseta de Kerguelen-oído.

Channichthys bospori (1995).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Se encuentra en el Océano Austral: región subantártica de la meseta de Kerguelen y el área de la barrera submarina Kerguelen-Heard.

Channichthys irinae (1995).- Es una especie de pez descrita por Shandikov, 1995. Es parte del género Channichthys y de la familia Channichthyidae. No hay subespecies enumeradas en el Catálogo de la vida. Los machos pueden alcanzar los 25,9 cm de longitud máxima. Esta especie es un consumidor típico de zooplancton. Se encuentra en el Océano Austral: isla Kerguelen

Channichthys mithridatis (2008) (Pez hielo verde).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Los machos pueden alcanzar los 37,7 cm de longitud máxima; las hembras 43,7 cm. Espinas dorsales 6-9; Radios blandos dorsales 32-34; Radios suaves anales 30-32; Vértebras 55-58. Esta especie se distingue por los siguientes caracteres: ancho interorbital estrecho, 6.3-7.7 veces en HL, 1.2-1.7 más corto que el diámetro de la órbita horizontal; ojos grandes, orbitan 4.5-5.6 veces en HL y 2.1-2.7 en la longitud del hocico; borde posterior del maxilar que se extiende por debajo de 1 / 2-2 / 3 del diámetro de la órbita; una hilera de rastrillos en la parte inferior del primer arco branquial en el lado externo del ceratobranquial; primera aleta dorsal muy alta, 2.9-4 veces en SL, con la segunda o tercera espina más larga; la membrana de la aleta dorsal primera no llega a las puntas de las espinas más largas; aletas dorsales bien separadas, el borde posterior de la membrana de la aleta D1 no alcanza la base del primer radio de D2; placas óseas redondeadas generalmente ausentes de la parte anterior de la línea lateral mediana; tuberculación en los frontales y en la espina dorsal anterior muy tenue o ausente, ausente en el maxilar, dentario y branquiostáticos. Las hembras maduran a aproximadamente 30-32 cm LT o 26-28 cm SL. Una especie piscívora; los contenidos del estómago contienen pescado. Se encuentra en la Antártida: isla Kerguelen. Atrapado por redes de arrastre de fondo en un área de fauna benténica bentónica

Channichthys normani (1996).- Es una especie de pez descrita por Balushkin, 1996. Channichthys normani es parte de la familia Channichthys y de la familia Channichthyidae. No hay subespecies enumeradas en el Catálogo de la vida

Channichthys panticapaei (1995).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Los machos pueden alcanzar los 40,2 cm de longitud máxima; las hembras 36,1 cm. Espinas dorsales (total): 6-8; Radios blandos dorsales (total): 32-34; Radios suaves anales: 29-32; Vértebras: 55-57. Se distingue por los siguientes caracteres: ancho interorbital ancho, 4.4-5.4 veces en HL, diámetro de órbita más ancho que horizontal; ojos de tamaño moderado, órbita 5.4-6.5 veces en HL, y 2.6-3.1 en la longitud del hocico y 1.1-1 / 4 en el ancho interorbital; borde posterior del maxilar que se extiende por debajo de 1/2 del diámetro de la órbita; dos hileras de rastrillos en la parte inferior del primer arco branquial en ceratobranquial e hipobranquial: 11-17 branquispinas en hilera anterior, 6-15 en hilera posterior, 18-31 en total; primera aleta dorsal alta, 3.2-5.1 veces en SL, con 1 a 3 espina más larga; la membrana de la aleta D1 no llega a las puntas de las espinas más largas; aletas dorsales bien separadas, el borde posterior de la membrana de la aleta D1 no alcanza la base del primer radio de D2; placas óseas redondeadas gruesas muy desarrolladas en la parte anterior de la línea lateral mediana; Tuberculación ósea fuerte en los frontales, espinas dorsal, branquiostáticos, radios pélvicos, maxilares y dentarios. Coloración gris oscuro a uniformemente negruzca. Madura a aproximadamente 30 cm LT (27 cm SL) y el desove probablemente tenga lugar en junio-julio. Una especie thalassobathyal y planctívora. El contenido estomacal contiene macroplancton, Parathemisto gaudichaudi y Thysanoessa macrura. Se encuentra en el Océano Austral, Subantártico: Islas Kerguelen. Atrapado por redes de arrastre de fondo.

Channichthys rhinoceratus (1844) - (Draco rinoceronte.).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Los machos pueden alcanzar los 60 cm de longitud máxima; longitud común 40; peso máximo publicado 500 gr. Vértebras 54 - 58. Se alimenta principalmente de peces y ocasionalmente de algas. Los adultos se encuentran desde la costa cercana a más de 750 m. Se encuentra en el Océano Austral: endémico de la meseta Kerguelen-Heard, Antártida.

Channichthys richardsoni (2011) (draco robusto).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Los machos pueden alcanzar los 37,4 cm de longitud máxima. Espinas dorsales 7-8; Radios blandos dorsales 31-34; Radios suaves anales 29-32. Esta especie se caracteriza por los siguientes D1 VII-VIII, D2 31-34; A 29-32; pectoral 19-20; placas tubulares óseas (escamas) de la línea lateral dorsal 61-78 y de la línea lateral mediana 9-23, placas óseas separadas redondeadas en la parte anterior de la línea lateral mediana 4-28, como regla, más de 10; cantidad de branquiespinas en el lado externo de la parte inferior del primer arco 6-15. Esta especie se distingue además por lo siguiente: anchura interorbital estrecha, 1.0-1.4 veces en el diámetro de la órbita horizontal, 5.8-7.5 en HL y 2.0-2.4 en altura de la cabeza a nivel del centro del ojo; ojo moderado en tamaño, su diámetro 5.2-6.3 veces en HL y 2.6-3.0 en la longitud del hocico; supraorbitales bordes óseos externos de los frontales notablemente elevados; hocico relativamente largo, aproximadamente igual o algo más corto que la mitad de la longitud de la cabeza, 2.1-2.2 en HL; las puntas de las mandíbulas están alineadas o la mandíbula inferior ligeramente protruida, los dientes en la sínfisis no visibles; borde posterior del maxilar que se extiende por debajo de 1 / 3-1 / 2 del diámetro de la órbita; una fila de rastrillos en la parte inferior del primer arco branquial en el lado externo del ceratobranquial; aleta pectoral que se extiende por encima del ano o el origen del primer radio de la aleta anal; primera aleta dorsal alta, 3.3-4.7 veces en SL, segundo y tercer radio más largo; la membrana de la aleta D1 no alcanza las puntas de los radios más largos, su altura es aproximadamente 1.4-1.7 veces la altura de D1; aletas dorsales bien separadas, el borde posterior de la membrana de la aleta D1 no alcanza la base del primer radio de D2; distancia interdorsal amplia, 10.9-16.4 veces en SL o 1.0-1.9 veces en longitud de D1; tuberculación ósea bien desarrollada en frontales y lagrimales, espinas flexibles D1, branquiostáticos, aletas pélvicas y en estructuras óseas de ambas líneas laterales; ausente en el maxilar y en la parte anterior de la mandíbula inferior.

Las hembras maduran a aproximadamente 29-31 cm LT (25-27 cm SL); el SGM VI-III posterior al desove se detecta claramente en hembras de más de 31 cm LT y el SGM III - en las más pequeñas, en primer lugar maduras. El SGM de los machos examinados varió de la etapa II a la etapa inicial III en la muestra de 32.5 cm LT. El SGM antes del desove y el engendro estaban ausentes. El período de desove aparentemente toma un lugar desde el otoño hasta el comienzo del invierno. Aparentemente un depredador principalmente piscívoro (inferido de la única fila de branquiespinas y similar a sus congéneres). La mayoría de los especímenes fueron capturados a una profundidad de 126 m en un área con abundante fauna bentónica y la alga Macrocystis pyrifera. Se encuentra en las aguas de plataforma de las islas y se captura en 2 redes de arrastre de fondo a profundidades de 126 y 270-310 m, junto con C. rhinoceratus , C. rugosus , C. panticapaei , C. bospori , C. irinae y C. mithridatis. Se encuentra en el Océano Austral, Subantártico: Islas Kerguelen.

Channichthys rugosus (1913).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Los machos pueden alcanzar los 3 cm de longitud máxima. Espinas anales: 0; Radios suaves anales: 28-29; Vértebras: 52. Se encuentra en el Océano Austral: Islas Kerguelen.

Channichthys velifer (1974).- Es un pez del género Channichthys . Se clasifica en la familia Fishichthyidae. Los machos pueden alcanzar los 49 cm de longitud máxima. La comida incluye cefalópodos y peces. Se encuentra en el Océano Austral: Subantártico, endémico de la meseta Kerguelén.

Género Chionobathyscus (1978).- Es un género monotipico de la familia Channichthyidae del orden de los Perciformes.

Chionobathyscus dewitti (1978).- Es una especie de draco rayado nativo del Océano Austral , donde ocurre a profundidades de 500 a 2,000 metros (1,600 a 6,600 pies). La especie, que crece hasta una longitud de 60 centímetros (24 pulgadas ) de LT , es el único miembro conocido de su género. Los adultos se alimentan principalmente de krill y peces. Se encuentra en la plataforma continental y la pendiente. Se encuentra en el Océano Austral: probablemente circum-Antártico en la plataforma continental y la pendiente.

Género Chionodraco (1906).- Es un género de la familia Channichthyidae del orden de los Perciformes nativos del Océano Austral. Al igual que muchos otros peces channichthyid , la reproducción en Chionodraco hamatus consiste en poner huevos en un nido y son protegidos por los padres. Las características morfológicas similares sugieren que al menos Chionodraco rastrospinosus tiene comportamientos reproductivos similares. Actualmente hay tres especies reconocidas en este género:

Chionodraco hamatus (1905).- Es un pez de la familia Channichthyidae. Las muestras varían de 33 a 37 cm, con una longitud máxima de 49 cm. Al igual que muchos otros peces channichthyid , la reproducción en Chionodraco hamatus consiste en poner huevos en un nido y proteger a los padres. Hombre prepara un nido y corteja a la hembra. La deposición de los huevos (y presumiblemente la fertilización también) tiene lugar en la noche. La hembra permanece en el nido, protegiendo y abanicando los huevos. La eclosión tiene lugar durante un período prolongado de tiempo. El hombre parece no estar involucrado en el cuidado de los huevos. Chionodraco hamatus se alimenta principalmente de peces pequeños, kril y larvas de peces. Esta especie vive en la zona demersal, y se puede encontrar en un rango de profundidad de 4 a 400 metros. Se encuentra en las frías aguas del Océano Austral en la plataforma continental que rodea la Antártida.

Chionodraco myersi (1960).- Es un pez de la familia Channichthyidae. Los machos pueden alcanzar los 38 cm de longitud. La fase pelágica larval es larga. Los adultos se alimentan de peces ( Pleuragramma antarcticum ) y eufáusidos ( Euphausia crystallorophias ). Este pez sólo se encuentra en los mares que circundan la Antártida, en la plataforma continental, entre los 200 y los 800 metros de profundidad. Se encuentra en el Océano Austral: Circun-Antártico en la plataforma continental.

Chionodraco rastrospinosus (1979) - Draco ocelado.- Es un pez de la familia Channichthyidae. Crecen hasta 52 centímetros (20 pulgadas) y un promedio de 30 centímetros (12 pulgadas). La hemoglobina le da a la sangre oxigenada su color rojo. A diferencia de otros vertebrados , los peces de la familia del draco rayado de la Antártida ( Channichthyidae ) no usan la hemoglobina para transportar oxígeno alrededor de sus cuerpos; en cambio, se utiliza la pequeña cantidad de oxígeno que simplemente se disuelve en el plasma sanguíneo. En 2011, Tokyo Sea Life Park afirmó que C. rastrospinosus tiene sangre totalmente transparente "como agua clara", después de diseccionar un espécimen. En 1954, Ruud notó que Chaenocephalus aceratus , otro miembro de esta familia, tenía sangre casi transparente, en contraste con la sangre amarillenta de otros miembros. C. aceratus y C. rastrospinosus no logran expresar la α-globina adulta mayor, α1, debido al mismo 5 'truncamiento del gen, y han perdido el gen de β-globina por completo. Zhao et al. Proponen que un ancestral channichthyid fish perdió la expresión de ambos genes a través de una sola mutación. El draco rayado antártico también tiene muy pocos eritrocitos . Se cree que se benefician de la pérdida de dependencia de los eritrocitos que contienen hemoglobina para el transporte de oxígeno al tener menos sangre viscosa y más fácilmente bombeable. Compensan esta pérdida al tener tasas metabólicas más bajas, branquias más grandes, piel sin escamas que pueden contribuir más al intercambio de gases, capilares más anchos y un aumento significativo en el volumen sanguíneo y el gasto cardíaco.

Las larvas tienen 17 mm de largo cuando nacen y crecen alrededor de 2 mm por semana. La etapa larval dura hasta 18 meses durante los cuales se alimentan principalmente de kril. Se vuelven sexualmente maduros a los cuatro años, y normalmente viven hasta ocho años, pero a veces hasta los doce. Tokyo Sea Life Park tiene el único pez cautivo. Un par de machos y hembras fueron donados junto con otras especies por los pescadores del arrastrero de krill Fukuei-maru como parte de un programa para recolectar capturas incidentales para el parque. En enero de 2013, la hembra engendró, y el 7 de mayo el primer huevo eclosionó con alrededor de 20 larvas más en las siguientes dos semanas. Anteriormente, especímenes vivos se han llevado a cabo para la investigación científica en otros lugares. Los adultos se alimentan de krill y otros peces. Vive en las frías aguas de la Antártida y es conocido por tener sangre libre de hemoglobina transparente. Se encuentra comúnmente en profundidades de 200 a 400 m. Se encuentra en el Océano Austral: varía desde las Islas Orcadas del Sur, las Islas Shetland del Sur hasta la Península Antártica.

Género Cryodraco (1900).- Es un género de la familia Channichthyidae del orden de los Perciformes nativos del Océano Austral. Actualmente hay tres especies reconocidas en este género:

Cryodraco antarcticus (1900).- Es un pez de la familia Channichthyidae. Los machos pueden alcanzar los 39,3 cm de longitud. Se presentan varias características biológicas de esta especie, con nueva información recopilada de estudios científicos recientes de las Islas Shetland del Sur y las Islas Orcadas del Sur. La especie está estrechamente relacionada con Chaenocephalus aceratus , pero se puede distinguir por una serie de caracteres merísticos. Las dos especies son similares en tamaño, coloración y forma del cuerpo, y tienen en común una serie de aspectos de su ciclo de vida, como la reproducción, la relación talla-peso y la dieta. Estas dos especies parecen ocupar un nicho muy similar en el ecosistema del arco del sur de Escocia. Sin embargo, Cryodraco antarcticus es menos abundante y utiliza diferentes componentes espaciales de las áreas de plataforma, reemplazando a Chaenocephalus aceratus en aguas más profundas y regiones de alta Antártica.

La fase pelágica larval es larga al final del invierno. Los adultos se alimentan de peces y krill. Es común en las aguas más profundas del arco del sur de Scotia y la zona alta antártica; rango de profundidad 90 - 600 m. Se encuentra en el Océano Austral: Islas Orcadas del Sur, Península Antártica y de los mares Weddell, Bellingshausen, Ross y Davis.

Cryodraco atkinsoni (1914).- Es un pez de la familia Channichthyidae. Los machos pueden alcanzar los 29,3 cm de longitud. Espinas dorsales 3; Radios blandos dorsales 42; Radios suaves anales: 46. Cabeza con manchas oscuras, tronco con barras; primer dorsal negruzco; aletas pélvicas oscuras. Se encuentra en el Océano Austral: Mar de Ross.

Cryodraco pappenheimi (1913).- Con base en un estudio morfométrico y merístico de 101 especímenes, reconocemos 2 especies válidas en el género de channichthyid antártico Cryodraco: Cryodraco antarcticus Dollo, 1900 y C. atkinsoni Regan, 1914. Aunque la especie se superpone en la mayoría de los caracteres merísticos y morfométricos, hemos distinguido varios caracteres confiables para el diagnóstico y la identificación. El escalado multidimensional, una técnica multivariante no paramétrica, separa claramente las dos especies en función de la longitud de la aleta pélvica, la longitud de la cabeza, el número de segundos radios de la aleta dorsal y el origen de la línea lateral inferior en relación con los radios de la aleta anal. Desde un punto de vista zoogeográfico, C. antarcticus tiene una distribución circumantártica, mientras que C. atkinsoni se limita en gran parte a la Provincia Zoogeográfica Antártica Oriental.

Género Dacodraco (1916).- Es un género monotipico de la familia Channichthyidae del orden de los Perciformes.

Dacodraco hunteri (1916).- Es una especie de draco rayado nativo del Océano Austral a profundidades de 300 a 800 metros (980 a 2.620 pies). Esta especie crece hasta una longitud de 29 centímetros (11 pulgadas ) de LT. La fase pelágica larval es larga. Se encuentra en el Océano Austral: Probablemente circum-Antártico en la plataforma continental. Esta especie es el único miembro conocido de su género

Género Neopagetopsis (1947).- Es un género monotipico de la familia Channichthyidae del orden de los Perciformes.

Neopagetopsis ionah (1947).- Es una especie de draco rayado de cocodrilo que se encuentra en el Océano Austral a profundidades de 20 a 900 metros (66 a 2,953 pies). Esta especie crece hasta una longitud de 56 centímetros (22 pulgadas ) de TL. Los adultos se alimentan principalmente de peces y kril. Se encuentra en el Océano Austral: Circun-Antártico en la plataforma continental y la pendiente. Esta especie es el único miembro conocido de su género

Género Pagetopsis (1913).- Es un género de pecescocodrilo encontrado en el Océano Austral. Actualmente hay dos especies reconocidas en este género:

Pagetopsis macropterus (1907).- Es un pez de la familia Channichthyidae. Los machos pueden alcanzar los 33 cm de longitud. Los adultos se alimentan principalmente de peces y krill. Se encuentra más comúnmente en profundidades de 5-40 m. Se encuentra en el Océano Austral: Circun-Antártico en la plataforma continental y las Islas Shetland del Sur.

Pagetopsis maculatus (1958).- Es un pez de la familia Channichthyidae. Los machos pueden alcanzar los 25 cm de longitud. La fase pelágica larval es larga. Se encuentra en el Océano Austral: Circun-Antártico en la plataforma continental.

Género Pseudochaenichthys (1937).- Es un género monotipico de la familia Channichthyidae del orden de los Perciformes.

Pseudochaenichthys georgianus (1937) - Draco cocodrilo.- Es una especie de draco rayado de cocodrilo que solo se conoce de la península Antártica septentrional y del mar de Escocia, donde se encuentra desde la superficie hasta 475 metros (1,558 pies). Esta especie crece hasta una longitud de 60 centímetros (24 pulgadas ) de TL. Longitud común 50 cm. Peso máximo publicado 2,1 kg. Los adultos se alimentan principalmente de krill y pescado. Generador síncrono. Se encuentra en el Océano Austral: conocido desde las islas del mar de Scotia y la parte norte de la Península Antártica hasta la isla de Scott. Es de menor importancia para las pesquerías comerciales . Esta especie es el único miembro conocido de su género
No hay comentarios:
Publicar un comentario